



Seth Blackshaw
@sethblackshaw.bsky.social
Professor of Neuroscience. Studying neural development, regeneration, and control of innate behaviors at Johns Hopkins.
@sethblackshaw at Twitter.
@sethblackshaw at Twitter.
Inspired by this, we tested rejuvenation strategies. We selectively expressed Yamanaka factors Oct4, Sox2, and Klf4 in Muller glia using a tamoxifen-inducible GlastCreER system with tetON control, driving low-level, sustained expression in adult mouse retina./29

September 12, 2025 at 8:41 PM
Inspired by this, we tested rejuvenation strategies. We selectively expressed Yamanaka factors Oct4, Sox2, and Klf4 in Muller glia using a tamoxifen-inducible GlastCreER system with tetON control, driving low-level, sustained expression in adult mouse retina./29
Using NicheNet, we identified rod-specific age-dependent niches: Niche 8, enriched in young rods with photoreceptor genes, and Niche 11, enriched in aged rods with Col4a3 and proinflammatory Ly75. These niches highlight distinct pro-youth vs. pro-aging states./28

September 12, 2025 at 8:38 PM
Using NicheNet, we identified rod-specific age-dependent niches: Niche 8, enriched in young rods with photoreceptor genes, and Niche 11, enriched in aged rods with Col4a3 and proinflammatory Ly75. These niches highlight distinct pro-youth vs. pro-aging states./28
Xenium spatial data also revealed some surprising aging-related cell-cell interactions. Muller glia exerted broad pro-rejuvenative effects on neighboring cells, especially rod photoreceptors, bipolar cells, and amacrine interneurons, suggesting they help buffer against aging-related decline./27

September 12, 2025 at 8:36 PM
Xenium spatial data also revealed some surprising aging-related cell-cell interactions. Muller glia exerted broad pro-rejuvenative effects on neighboring cells, especially rod photoreceptors, bipolar cells, and amacrine interneurons, suggesting they help buffer against aging-related decline./27
Gene expression changes observed using snRNA-Seq generally replicated in Xenium analysis. This shows plots of several aging-regulated genes in Muller glia./27

September 12, 2025 at 8:34 PM
Gene expression changes observed using snRNA-Seq generally replicated in Xenium analysis. This shows plots of several aging-regulated genes in Muller glia./27
To further examine aging-dependent changes in situ, we performed Xenium5k analysis in mouse retina, simultaneously measuring ~5,000 genes at 1 μm resolution. This mapped all major retinal cell types and enabled construction of cell-type-specific aging clocks across the tissue./26

September 12, 2025 at 8:30 PM
To further examine aging-dependent changes in situ, we performed Xenium5k analysis in mouse retina, simultaneously measuring ~5,000 genes at 1 μm resolution. This mapped all major retinal cell types and enabled construction of cell-type-specific aging clocks across the tissue./26
But as with gene expression and regulation, we detected few evolutionarily conserved patterns of cell-cell signaling./25

September 12, 2025 at 8:26 PM
But as with gene expression and regulation, we detected few evolutionarily conserved patterns of cell-cell signaling./25
In all three species, aged cells showed more complex patterns of cell-cell signaling, with Muller glia acting as signaling hubs, particularly in zebrafish and mouse./24

September 12, 2025 at 8:25 PM
In all three species, aged cells showed more complex patterns of cell-cell signaling, with Muller glia acting as signaling hubs, particularly in zebrafish and mouse./24
By integrating scRNA-Seq with scATAC-Seq, we mapped transcription factor activity in young vs. aged cells. This revealed robust age-dependent regulatory shifts, though again with strong dependence on species and cell type, and limited evolutionary conservation./20

September 12, 2025 at 8:20 PM
By integrating scRNA-Seq with scATAC-Seq, we mapped transcription factor activity in young vs. aged cells. This revealed robust age-dependent regulatory shifts, though again with strong dependence on species and cell type, and limited evolutionary conservation./20
Still, a few broad regulators emerged: fat4, an atypical cadherin, was age-regulated in zebrafish; Stat1, an inflammatory signal transducer, in mice; and metallothionein 1F, a stress response gene, in humans. These may represent conserved functional stress axes./19

September 12, 2025 at 8:14 PM
Still, a few broad regulators emerged: fat4, an atypical cadherin, was age-regulated in zebrafish; Stat1, an inflammatory signal transducer, in mice; and metallothionein 1F, a stress response gene, in humans. These may represent conserved functional stress axes./19
Indeed, only a tiny handful of genes displayed consistent aging-related changes across zebrafish, mice, and humans. Most genes altered in one species showed little or no regulation in the others, underscoring evolutionary divergence in retinal aging mechanisms./16

September 12, 2025 at 8:10 PM
Indeed, only a tiny handful of genes displayed consistent aging-related changes across zebrafish, mice, and humans. Most genes altered in one species showed little or no regulation in the others, underscoring evolutionary divergence in retinal aging mechanisms./16
Species, sex, and cell-type differences stand out most. For example, retinal ganglion cells (RGCs) in human females showed the largest number of age-regulated genes. Yet in mice and zebrafish, RGCs showed mid-range numbers, and in human males, changes were minimal./13

September 12, 2025 at 7:30 PM
Species, sex, and cell-type differences stand out most. For example, retinal ganglion cells (RGCs) in human females showed the largest number of age-regulated genes. Yet in mice and zebrafish, RGCs showed mid-range numbers, and in human males, changes were minimal./13
These three broad classes capture general expression trends: genes enriched in young cells, genes showing U- or inverted U-shaped midlife dynamics, and genes elevated in aged cells. While broadly similar across species, details varied strongly by sex and cell type./11

September 12, 2025 at 7:26 PM
These three broad classes capture general expression trends: genes enriched in young cells, genes showing U- or inverted U-shaped midlife dynamics, and genes elevated in aged cells. While broadly similar across species, details varied strongly by sex and cell type./11
And as has been reported for DNA methylation-based markers of cell age, we find that acute retina injury induced by either NMDA excitotoxicity or light damage induces a substantial though mostly transient increase in cellular age across multiple cell types./10

September 12, 2025 at 7:22 PM
And as has been reported for DNA methylation-based markers of cell age, we find that acute retina injury induced by either NMDA excitotoxicity or light damage induces a substantial though mostly transient increase in cellular age across multiple cell types./10
Using elastic net regression analysis, we were then able to identify gene sets that serve as highly predictive aging clocks for all retinal cell types in each species./9

September 12, 2025 at 7:19 PM
Using elastic net regression analysis, we were then able to identify gene sets that serve as highly predictive aging clocks for all retinal cell types in each species./9
We next clustered genes showing age-dependent expression changes across species. This analysis revealed ~10–12 major clusters, each representing a distinct temporal pattern of regulation, ranging from youth-biased to middle-aged peaks to late-life upregulation./8
September 12, 2025 at 7:15 PM
We next clustered genes showing age-dependent expression changes across species. This analysis revealed ~10–12 major clusters, each representing a distinct temporal pattern of regulation, ranging from youth-biased to middle-aged peaks to late-life upregulation./8
To tackle this, we conducted a particularly large-scale comparative multiomic analysis across the natural lifespan. In zebrafish and mice, we analyzed ~200,000 cells across 10 timepoints. Human samples, generated by Rui Chen’s Baylor group, add >2 million cells, skewing toward older ages./7

September 12, 2025 at 7:14 PM
To tackle this, we conducted a particularly large-scale comparative multiomic analysis across the natural lifespan. In zebrafish and mice, we analyzed ~200,000 cells across 10 timepoints. Human samples, generated by Rui Chen’s Baylor group, add >2 million cells, skewing toward older ages./7
Like development and acute injury, aging involves dynamic changes in gene expression over time. But unlike those processes, it is much harder to study: changes unfold slowly, are less robust, and far more variable across cells, individuals, and environmental contexts./3

September 12, 2025 at 7:06 PM
Like development and acute injury, aging involves dynamic changes in gene expression over time. But unlike those processes, it is much harder to study: changes unfold slowly, are less robust, and far more variable across cells, individuals, and environmental contexts./3
Aging is a primary risk factor in the development of polygenic blinding diseases such as AMD and glaucoma, as shown in these graphs. This raises the question of whether it might be possible to treat these disorders at least in part by targeting the underlying process of cellular aging./2

September 12, 2025 at 7:05 PM
Aging is a primary risk factor in the development of polygenic blinding diseases such as AMD and glaucoma, as shown in these graphs. This raises the question of whether it might be possible to treat these disorders at least in part by targeting the underlying process of cellular aging./2
Interestingly, we had previously shown that preventing Lhx6+ cells from being formed in the first place showed an opposite effect on sleep homeostasis, with mutants sleeping less. This indicates that Lhx6+ ZI neurons play a central and complex role in both regulating sleep pressure and sleep need./7

August 5, 2025 at 2:26 PM
Interestingly, we had previously shown that preventing Lhx6+ cells from being formed in the first place showed an opposite effect on sleep homeostasis, with mutants sleeping less. This indicates that Lhx6+ ZI neurons play a central and complex role in both regulating sleep pressure and sleep need./7
We have that high sleep pressure activates Lhx6 ZI neurons, that they induce sleep with highly delayed kinetics, and that they form a node of a neural circuit that senses experimentally-induced sleep pressure with glutamatergic neurons of the thalamic reuniens nucleus./2

August 5, 2025 at 2:10 PM
We have that high sleep pressure activates Lhx6 ZI neurons, that they induce sleep with highly delayed kinetics, and that they form a node of a neural circuit that senses experimentally-induced sleep pressure with glutamatergic neurons of the thalamic reuniens nucleus./2
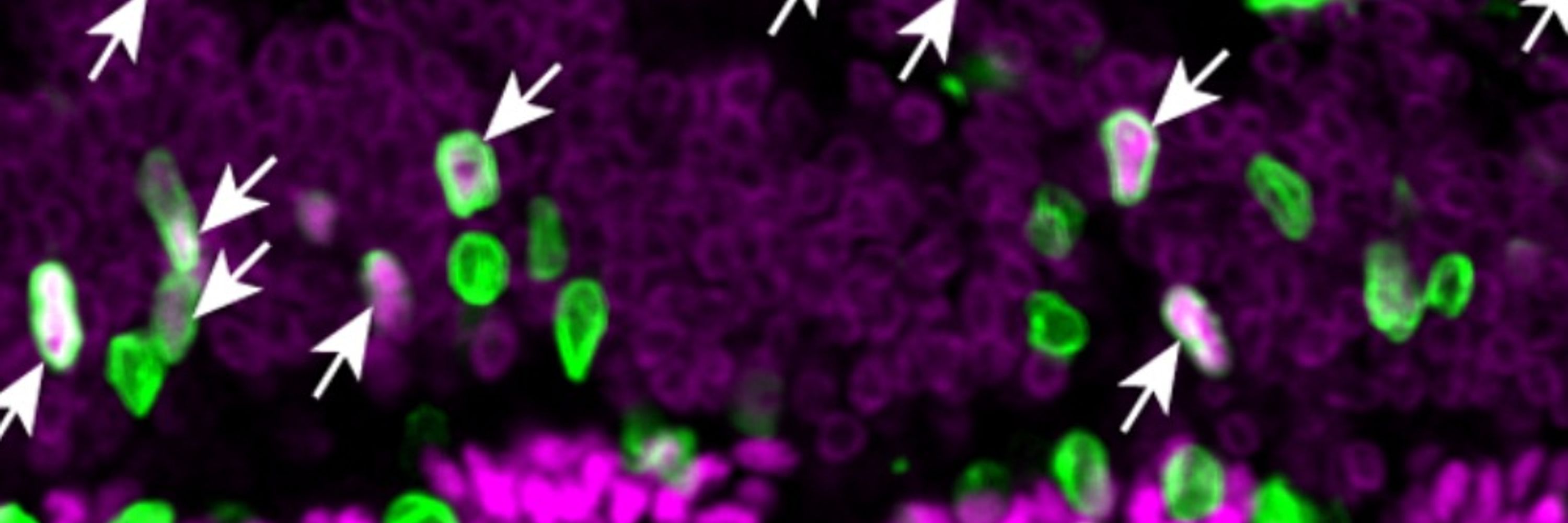
Clearly there will other neuronal populations that relay this information to the Lhx6 ZI neurons, although since these cells receive direct projections from dozens of brain regions, there are many potential candidates./18
June 19, 2025 at 7:31 PM
Clearly there will other neuronal populations that relay this information to the Lhx6 ZI neurons, although since these cells receive direct projections from dozens of brain regions, there are many potential candidates./18
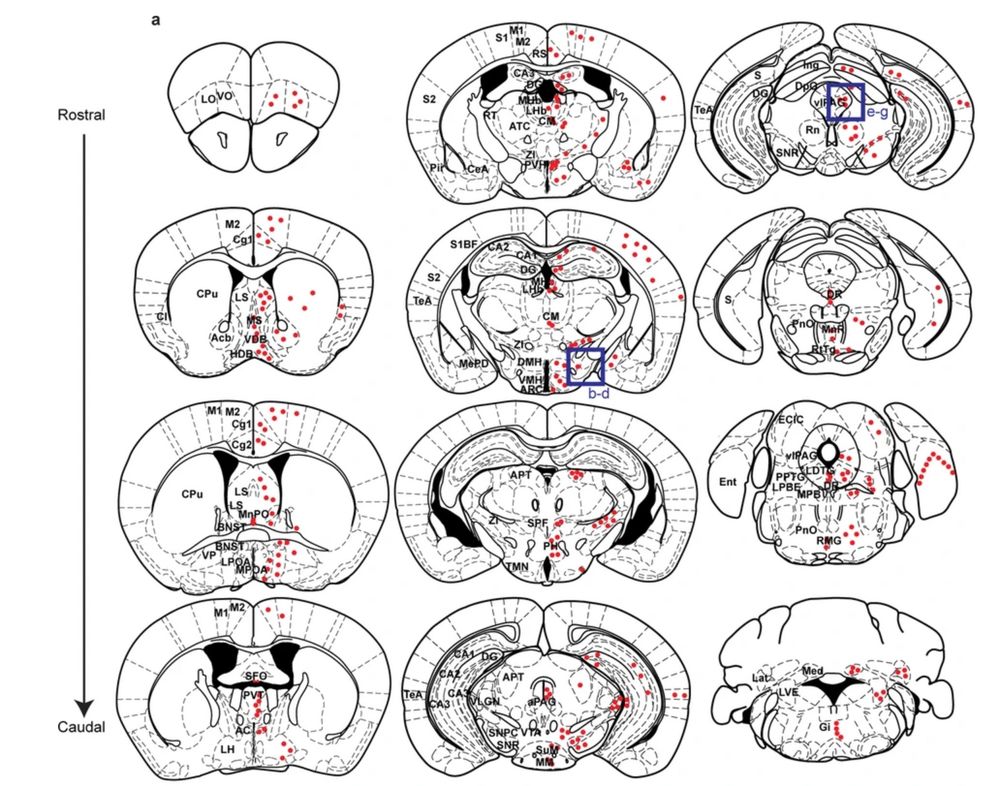
This work establishes the ReN-Lhx6 ZI circuit as essential for both sensing and signaling levels of homeostatic sleep pressure, but leaves many questions unanswered./15
June 19, 2025 at 7:23 PM
This work establishes the ReN-Lhx6 ZI circuit as essential for both sensing and signaling levels of homeostatic sleep pressure, but leaves many questions unanswered./15
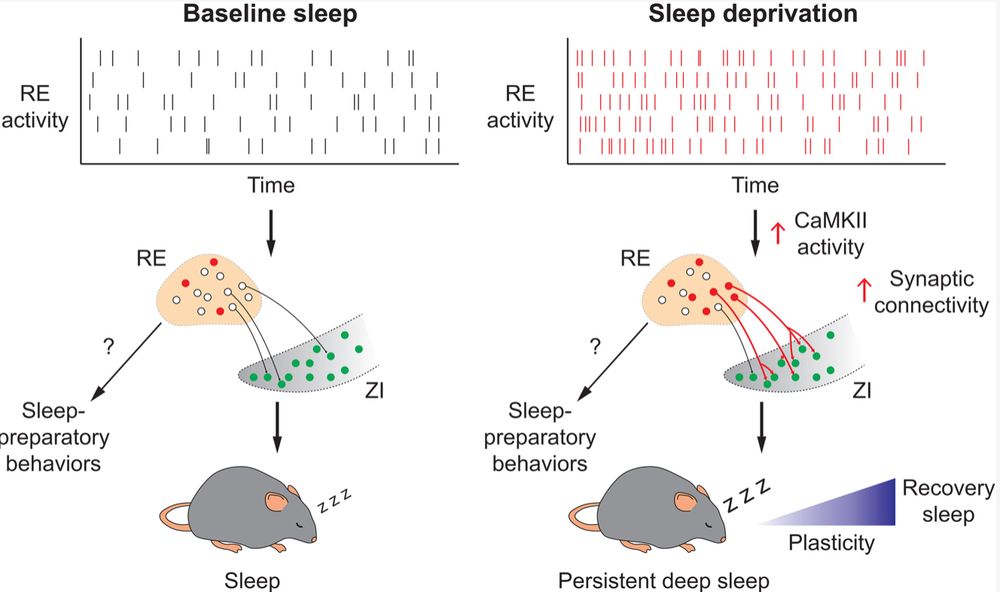
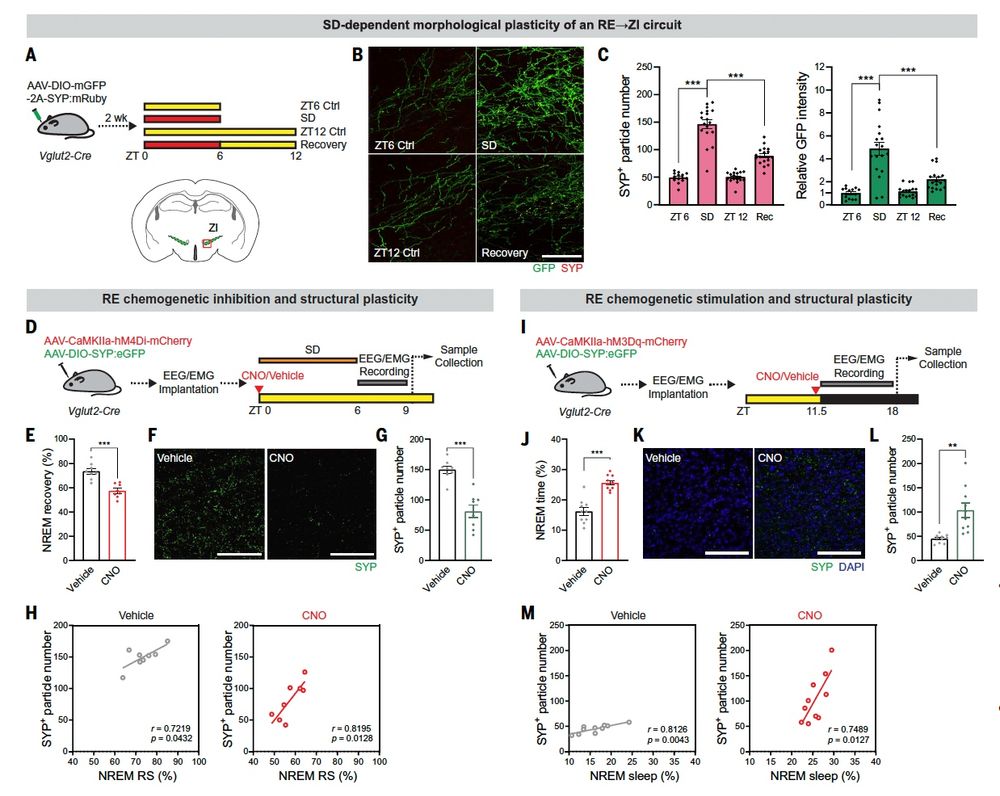
What's more, synaptic connections between ReN and Lhx6 ZI neurons are both physically and functionally strengthened by sleep deprivation in a CamKII-dependent manner, and these are in turn reversed by recovery sleep./14
June 19, 2025 at 7:20 PM
What's more, synaptic connections between ReN and Lhx6 ZI neurons are both physically and functionally strengthened by sleep deprivation in a CamKII-dependent manner, and these are in turn reversed by recovery sleep./14
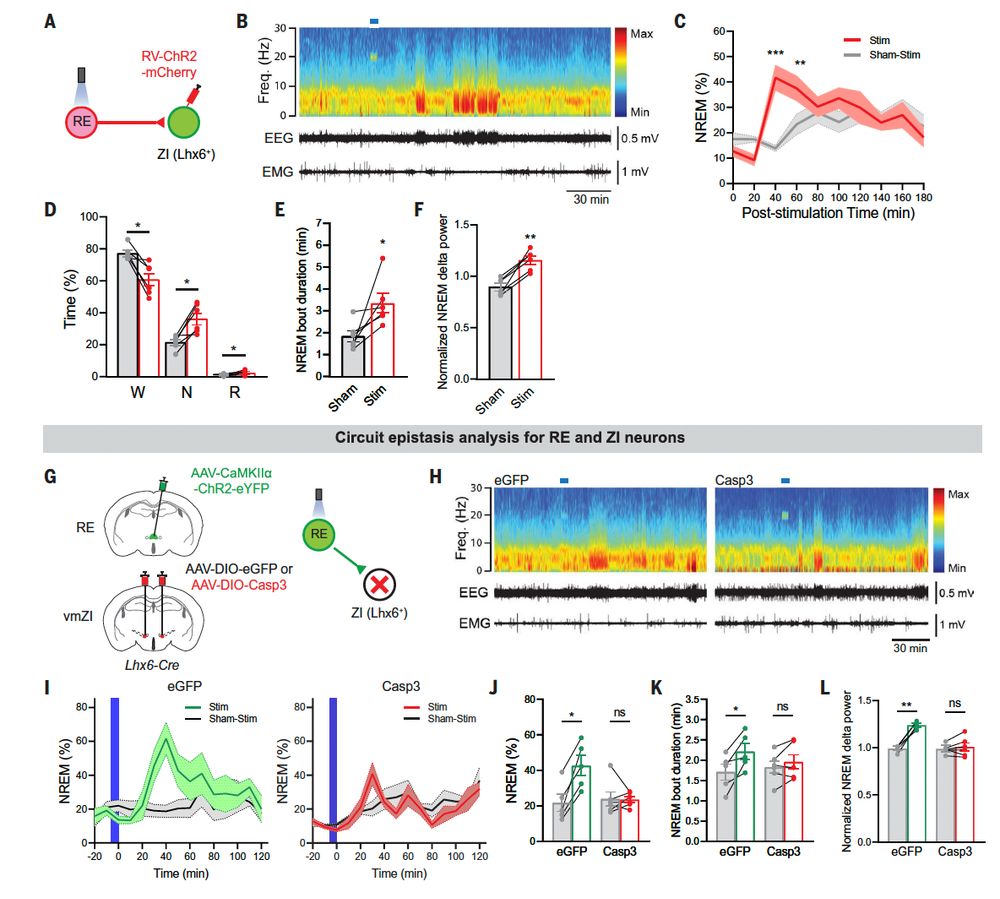
So where do the Lhx6-positive cells fit in? It turns out that if you selectively activate ReN projections to the ZI you phenocopy the effects of whole cell activation, and if you kill the Lhx6-positive neurons you no longer observe ReN-induced sleep./13
June 19, 2025 at 7:18 PM
So where do the Lhx6-positive cells fit in? It turns out that if you selectively activate ReN projections to the ZI you phenocopy the effects of whole cell activation, and if you kill the Lhx6-positive neurons you no longer observe ReN-induced sleep./13
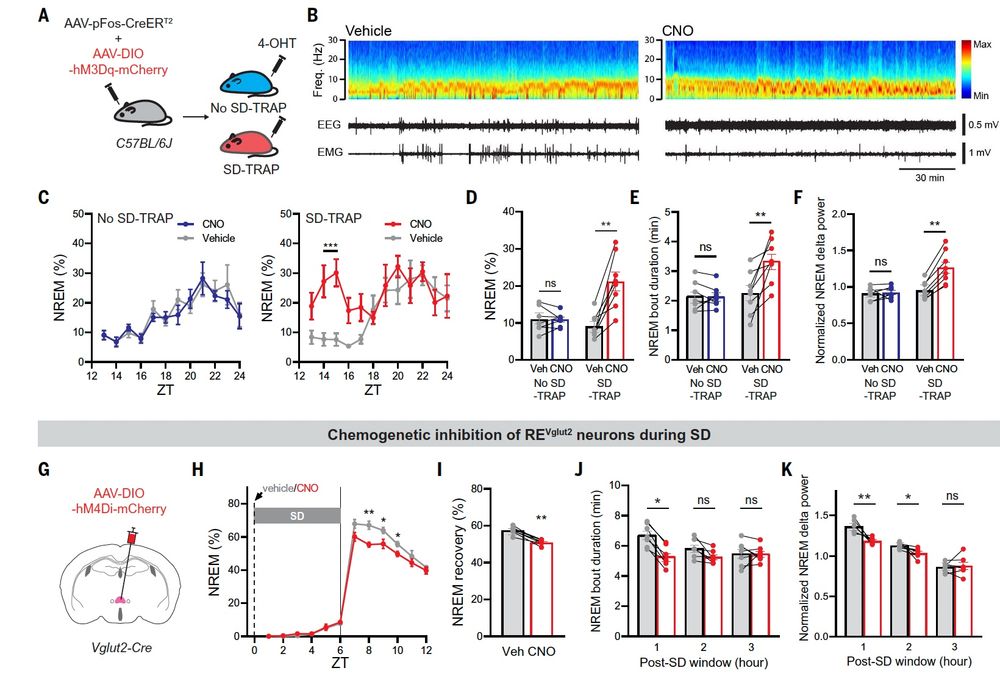
ReN neurons are activated during sleep deprivation and silenced during recovery sleep, and are themselves necessary for recovery sleep.12
June 19, 2025 at 7:15 PM
ReN neurons are activated during sleep deprivation and silenced during recovery sleep, and are themselves necessary for recovery sleep.12