




Marcin J. Suskiewicz
@msuskiewicz.bsky.social
Structural biologist and biochemist. CNRS researcher at CBM Orléans @cbm-upr4301.bsky.social. Interested in protein modifications & interactions. Also husband, dad of 2, friend, ☧. Personal website: msuskiewicz.github.io
We suggest defining four hydrophobic subpockets (HS1-HS4) on SUMO, each recognising one residue of the hydrophobic SIM core. In addition, we define the β strand-binding subpocket (βS) for binding the β-strand part of SIM, and the phospho/acidic-binding subpocket (PS) interacting with acidic part.

November 24, 2025 at 9:12 PM
We suggest defining four hydrophobic subpockets (HS1-HS4) on SUMO, each recognising one residue of the hydrophobic SIM core. In addition, we define the β strand-binding subpocket (βS) for binding the β-strand part of SIM, and the phospho/acidic-binding subpocket (PS) interacting with acidic part.
Our focus is on structural and evolutionary aspects. Which SUMO surfaces and residues mediate its different interactions? How is SUMO distinguished from ubiquitin to create a specific, orthogonal PTM system? What do the surface properties of SUMO imply for the evolution of its binders and function?

November 5, 2025 at 1:49 PM
Our focus is on structural and evolutionary aspects. Which SUMO surfaces and residues mediate its different interactions? How is SUMO distinguished from ubiquitin to create a specific, orthogonal PTM system? What do the surface properties of SUMO imply for the evolution of its binders and function?
A part of our team at the recent EMBO SUMO conference in La Grande Motte/Montpellier

September 30, 2025 at 9:43 AM
A part of our team at the recent EMBO SUMO conference in La Grande Motte/Montpellier
Very happy that our SIAH1 article - published online in February - was now chosen for the cover of the new issue of the @febsj.bsky.social . Thank you @spartelab.bsky.social for the wonderful collaboration and the lovely cellular images. febs.onlinelibrary.wiley.com/toc/17424658...

June 5, 2025 at 5:49 AM
Very happy that our SIAH1 article - published online in February - was now chosen for the cover of the new issue of the @febsj.bsky.social . Thank you @spartelab.bsky.social for the wonderful collaboration and the lovely cellular images. febs.onlinelibrary.wiley.com/toc/17424658...
PhD and postdoc positions with a brilliant friend of mine, Martin Lukačišin in Bratislava, Slovakia. Check out his website and papers lukacisinlab.github.io

April 28, 2025 at 12:30 PM
PhD and postdoc positions with a brilliant friend of mine, Martin Lukačišin in Bratislava, Slovakia. Check out his website and papers lukacisinlab.github.io
@clausenlab.bsky.social Greetings from the beamline!

February 6, 2025 at 7:19 PM
@clausenlab.bsky.social Greetings from the beamline!
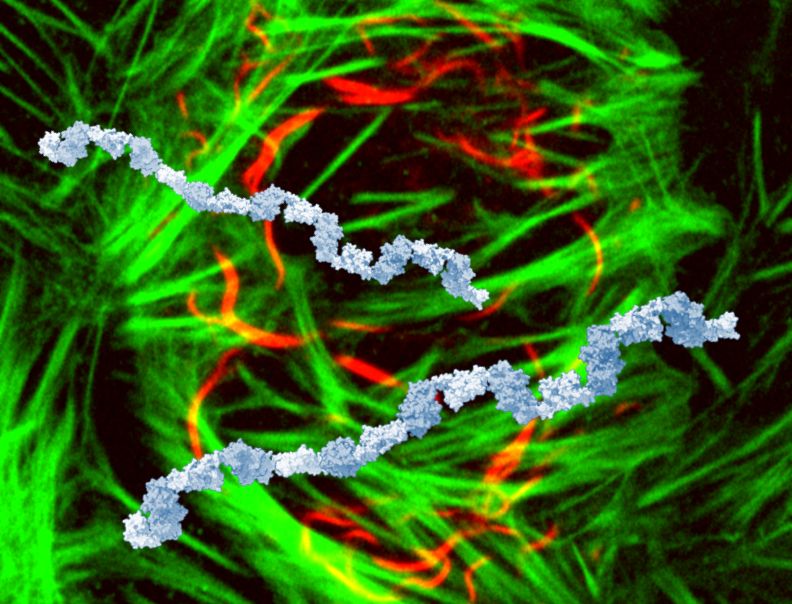
This is a cover design that wasn't accepted, showing SIAH1 multimers on top of cells (SIAH1 red, actin green). We were not able to visualise SIAH1 chains in vitro, as we could only purify N- or C-term halves of SIAH1 separately: the blue chains are made from combined dimeric structures of each half.
February 6, 2025 at 10:17 AM
This is a cover design that wasn't accepted, showing SIAH1 multimers on top of cells (SIAH1 red, actin green). We were not able to visualise SIAH1 chains in vitro, as we could only purify N- or C-term halves of SIAH1 separately: the blue chains are made from combined dimeric structures of each half.
Lastly, Lucija performed nice in-vitro ubiquitylation assays showing that the V90R mutant of the RING-ZnF1 fragment is deficient in making long ubiquitin chains. So there is also a role for RING dimerisation in catalysing polyubiquitylation, as seen for other dimeric RING Ub E3s.

February 6, 2025 at 10:05 AM
Lastly, Lucija performed nice in-vitro ubiquitylation assays showing that the V90R mutant of the RING-ZnF1 fragment is deficient in making long ubiquitin chains. So there is also a role for RING dimerisation in catalysing polyubiquitylation, as seen for other dimeric RING Ub E3s.
When we look at the colocalisation with an aggregated substrate, Synphilin-1, SIAH1 WT does colocalise, but this is largely prevented when we block RING domain dimerisation with the V90R mutation. This leads us to a model where multimerisation allows high-avidity binding to multivalent substrates.


February 6, 2025 at 10:01 AM
When we look at the colocalisation with an aggregated substrate, Synphilin-1, SIAH1 WT does colocalise, but this is largely prevented when we block RING domain dimerisation with the V90R mutation. This leads us to a model where multimerisation allows high-avidity binding to multivalent substrates.
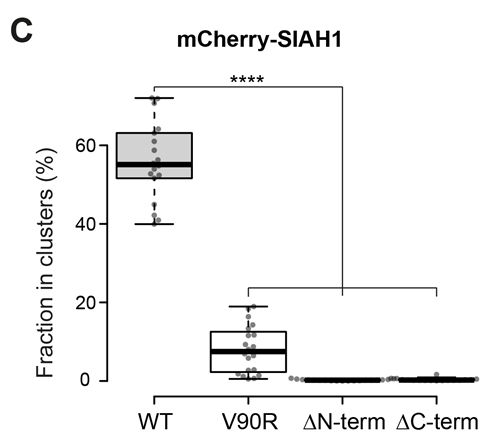
Here's what happens in cells: mCherry-tagged SIAH1 (admittedly, somewhat overexpressed) localises to elongated clusters, but you need to have both RING and SBD for that, and no V90R mutation, otherwise SIAH1 is mostly diffuse.

February 6, 2025 at 9:58 AM
Here's what happens in cells: mCherry-tagged SIAH1 (admittedly, somewhat overexpressed) localises to elongated clusters, but you need to have both RING and SBD for that, and no V90R mutation, otherwise SIAH1 is mostly diffuse.
We crystallised & Franck solved the crystal structure of the dimer formed by the RING-ZnF1 part of SIAH1. We confirmed dimerisation in solution, & showed V90R mutation blocks RING dimerisation. @origichals.bsky.social did lovely AlphaFold modelling showing dimerisation across the SINA/SIAH family.

February 6, 2025 at 9:54 AM
We crystallised & Franck solved the crystal structure of the dimer formed by the RING-ZnF1 part of SIAH1. We confirmed dimerisation in solution, & showed V90R mutation blocks RING dimerisation. @origichals.bsky.social did lovely AlphaFold modelling showing dimerisation across the SINA/SIAH family.
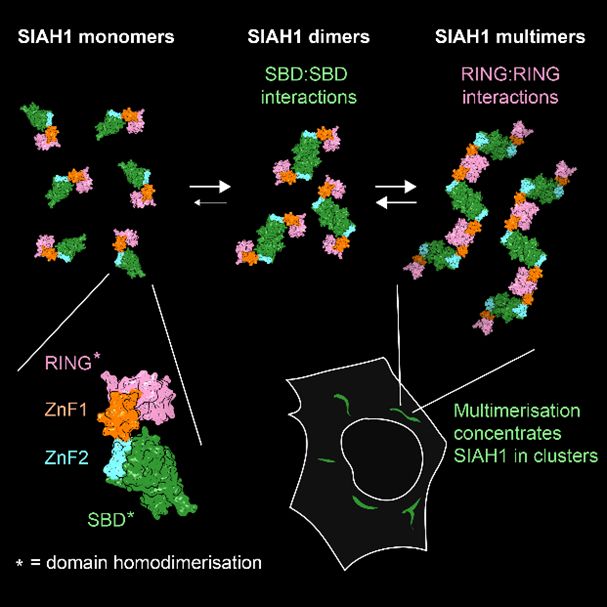
Happy to share our new study showing that the Ub E3 ligase SIAH1, known to dimerise via its C-term SBD, also dimerises via its N-term RING. When these tendencies combine, full-length SIAH1 forms multimers, which might explain its clustering in cells & preference for aggregated/multimeric substrates.
February 6, 2025 at 9:29 AM
Happy to share our new study showing that the Ub E3 ligase SIAH1, known to dimerise via its C-term SBD, also dimerises via its N-term RING. When these tendencies combine, full-length SIAH1 forms multimers, which might explain its clustering in cells & preference for aggregated/multimeric substrates.
We are still considering postdoc candidates for the post advertised below. A 2-year contract beginning in 2025 to work on protein SUMO E3 ligases and the nucleosome.

November 16, 2024 at 6:43 AM
We are still considering postdoc candidates for the post advertised below. A 2-year contract beginning in 2025 to work on protein SUMO E3 ligases and the nucleosome.
A recent group photo on the banks of the Loiret ☀️

November 14, 2024 at 6:34 AM
A recent group photo on the banks of the Loiret ☀️