




Ulrich Hohmann
@hohmannulrich.bsky.social
Finally, we test the linearity of this model through IP-MS of UAP56 mutants. If we perturb TREX-2 binding (UAP56 mutant M1) -> UAP56 accumulates on mRNPs. THO binding (M2) -> UAP56 depletes from mRNPs. THO+TREX-2 binding (M3) -> mirrors loss of THO binding alone. 17/

November 19, 2025 at 11:22 PM
Finally, we test the linearity of this model through IP-MS of UAP56 mutants. If we perturb TREX-2 binding (UAP56 mutant M1) -> UAP56 accumulates on mRNPs. THO binding (M2) -> UAP56 depletes from mRNPs. THO+TREX-2 binding (M3) -> mirrors loss of THO binding alone. 17/
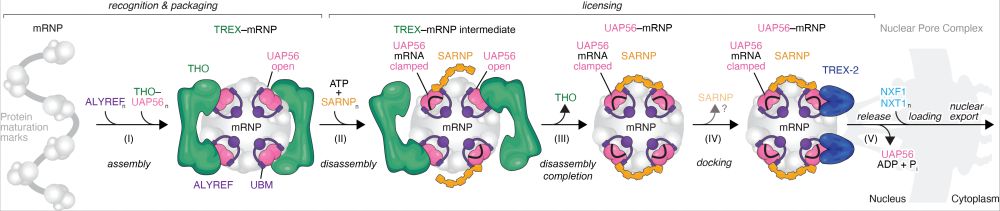
Taken together, we find that UAP56 is an “mRNP packaging mark”, that controls important aspects of the nuclear events of the mRNA export pathway: disassembly of TREX, docking of the mRNP at the NPC, and release into the NPC. 16/
November 19, 2025 at 11:22 PM
Taken together, we find that UAP56 is an “mRNP packaging mark”, that controls important aspects of the nuclear events of the mRNA export pathway: disassembly of TREX, docking of the mRNP at the NPC, and release into the NPC. 16/
We can visualize this TREX-2 activity also directly: we bead-immobilize UAP56–ADP-Pi–RNA complexes. Incubation with TREX-2 releases UAP56 from the RNA. In contrast, as expected, the THO complex does not have any effect on clamped UAP56. 15/

November 19, 2025 at 11:22 PM
We can visualize this TREX-2 activity also directly: we bead-immobilize UAP56–ADP-Pi–RNA complexes. Incubation with TREX-2 releases UAP56 from the RNA. In contrast, as expected, the THO complex does not have any effect on clamped UAP56. 15/
A highly conserved ‘wedge’ loop of the TREX-2 subunit GANP binds the nucleotide, which is still bound to UAP56’s RecA1 lobe (unusual for an open DExD ATPase). In vitro ATPase assays show a dramatic stimulation of UAP56’s ATPase rate by TREX-2. So TREX-2 promotes RNA release from UAP56! 14/

November 19, 2025 at 11:22 PM
A highly conserved ‘wedge’ loop of the TREX-2 subunit GANP binds the nucleotide, which is still bound to UAP56’s RecA1 lobe (unusual for an open DExD ATPase). In vitro ATPase assays show a dramatic stimulation of UAP56’s ATPase rate by TREX-2. So TREX-2 promotes RNA release from UAP56! 14/
In cells we show: the identified interfaces are critical for cell viability. In addition, perturbing the UAP56–TREX-2 interaction leads to mRNA export defects, as assessed (amongst other assays) through polyA-FISH in human K562 cells with a knockout-rescue setup for the TREX-2 subunit PCID2. 12/

November 19, 2025 at 11:22 PM
In cells we show: the identified interfaces are critical for cell viability. In addition, perturbing the UAP56–TREX-2 interaction leads to mRNA export defects, as assessed (amongst other assays) through polyA-FISH in human K562 cells with a knockout-rescue setup for the TREX-2 subunit PCID2. 12/
Could TREX-2 be a landing pad at the NPC for UAP56-bound, export ready mRNPs? Biochemically we show that UAP56 does indeed bind TREX-2 and solved a cryo-EM strcture of the complex. This revealed a role for the conserved UAP56 NTD in TREX-2 binding. 11/

November 19, 2025 at 11:22 PM
Could TREX-2 be a landing pad at the NPC for UAP56-bound, export ready mRNPs? Biochemically we show that UAP56 does indeed bind TREX-2 and solved a cryo-EM strcture of the complex. This revealed a role for the conserved UAP56 NTD in TREX-2 binding. 11/
Besides known UAP56 interactors we find some novel ones. Among them: GANP and PCID2, two subunits of the TREX-2 complex. A big surprise to us! TREX-2 (sharing only its name with TREX) is involved in mRNA export and bound to the basket of the nuclear pore complex (NPC)… 10/

November 19, 2025 at 11:22 PM
Besides known UAP56 interactors we find some novel ones. Among them: GANP and PCID2, two subunits of the TREX-2 complex. A big surprise to us! TREX-2 (sharing only its name with TREX) is involved in mRNA export and bound to the basket of the nuclear pore complex (NPC)… 10/
Remarkably, SARNP disassembles THO-UAP56 in the presence of ATP and RNA in vitro. And in endogenous mRNPs? We purified endogenous TREX-bound mRNPs. Again, a minimal SARNP construct (including the UCM) could dissociate the mRNP from THO. 8/

November 19, 2025 at 11:22 PM
Remarkably, SARNP disassembles THO-UAP56 in the presence of ATP and RNA in vitro. And in endogenous mRNPs? We purified endogenous TREX-bound mRNPs. Again, a minimal SARNP construct (including the UCM) could dissociate the mRNP from THO. 8/
Is RNA clamping of UAP56 regulated? We identify SARNP as a ‘clamping factor’. It has a motif we named ‘UCM’, which binds adjacent to the newly identified N-UBM binding site on UAP56. N-UBM and UCM cooperatively enhance UAP56’s affinity for RNA, this might aid directionality of mRNP remodelling. 7/

November 19, 2025 at 11:22 PM
Is RNA clamping of UAP56 regulated? We identify SARNP as a ‘clamping factor’. It has a motif we named ‘UCM’, which binds adjacent to the newly identified N-UBM binding site on UAP56. N-UBM and UCM cooperatively enhance UAP56’s affinity for RNA, this might aid directionality of mRNP remodelling. 7/
Upon RNA clamping UAP56 hydrolyses ATP, and the resulting UAP56–ADP-Pi–RNA complexes are remarkably stable, for example persisting in size exclusion chromatography experiments. This suggests that UAP56 can be ‘deposited’ on the mRNA as a maturation mark. 6/

November 19, 2025 at 11:22 PM
Upon RNA clamping UAP56 hydrolyses ATP, and the resulting UAP56–ADP-Pi–RNA complexes are remarkably stable, for example persisting in size exclusion chromatography experiments. This suggests that UAP56 can be ‘deposited’ on the mRNA as a maturation mark. 6/
Could the conformational change in UAP56 between its open and its RNA-clamped state play a role? Turns out it does: Open UAP56 binds THO, RNA-clamped UAP56 has no measurable affinity! This suggests that clamping of UAP56 releases THO from the mRNP. 5/

November 19, 2025 at 11:22 PM
Could the conformational change in UAP56 between its open and its RNA-clamped state play a role? Turns out it does: Open UAP56 binds THO, RNA-clamped UAP56 has no measurable affinity! This suggests that clamping of UAP56 releases THO from the mRNP. 5/
We found that UAP56 binds to the two known UAP56-binding motifs (UBMs) in mRNP export adapters, such as ALYREF, in distinct ways. The C-UBM binds the previously known site on UAP56’s RecA1 lobe, but, unexpectedly, the N-UBM binds a distinct site on the RecA2 lobe. 4/

November 19, 2025 at 11:22 PM
We found that UAP56 binds to the two known UAP56-binding motifs (UBMs) in mRNP export adapters, such as ALYREF, in distinct ways. The C-UBM binds the previously known site on UAP56’s RecA1 lobe, but, unexpectedly, the N-UBM binds a distinct site on the RecA2 lobe. 4/
THO binds to maturing mRNPs (=mRNA+proteins), directing them into the mRNA export pathway. Yet THO must again dissociate, to allow for subsequent steps in export. How? The connection between mRNP and THO is the DExD-box ATPase UAP56, which binds to UBM motifs in mRNP export adapters (like ALYREF).3/

November 19, 2025 at 11:22 PM
THO binds to maturing mRNPs (=mRNA+proteins), directing them into the mRNA export pathway. Yet THO must again dissociate, to allow for subsequent steps in export. How? The connection between mRNP and THO is the DExD-box ATPase UAP56, which binds to UBM motifs in mRNP export adapters (like ALYREF).3/
Finally out in @nature.com! We uncovered a mechanistic framework for a general and conserved mRNA nuclear export pathway. www.nature.com/articles/s41.... 1/
November 19, 2025 at 11:22 PM
Finally out in @nature.com! We uncovered a mechanistic framework for a general and conserved mRNA nuclear export pathway. www.nature.com/articles/s41.... 1/
In sum our work reveals that (m)RNA-clamped UAP56 not only promotes mRNA export by binding TREX-2 at the NPC. It can also be read out by the LENG8-PS module in PAXT to destine the RNA for decay. (9/x)

September 22, 2025 at 11:23 PM
In sum our work reveals that (m)RNA-clamped UAP56 not only promotes mRNA export by binding TREX-2 at the NPC. It can also be read out by the LENG8-PS module in PAXT to destine the RNA for decay. (9/x)
Perturbation of the LENG8–ZFC3H1 interface, or rapid depletion of either protein, leads to the upregulation of bona fide PAXT polyA-RNA targets. This demonstrates that the LENG8-PS module is functionally important for PAXT function. (8/x)

September 22, 2025 at 11:23 PM
Perturbation of the LENG8–ZFC3H1 interface, or rapid depletion of either protein, leads to the upregulation of bona fide PAXT polyA-RNA targets. This demonstrates that the LENG8-PS module is functionally important for PAXT function. (8/x)
Alphafold reveals a conserved mode of interaction between LENG8 and the PAXT scaffolding subunit ZFC3H1, which we can validate in vivo. Thus, LENG8-PS constitutes a TREX-2-like module of PAXT. (7/x)

September 22, 2025 at 11:23 PM
Alphafold reveals a conserved mode of interaction between LENG8 and the PAXT scaffolding subunit ZFC3H1, which we can validate in vivo. Thus, LENG8-PS constitutes a TREX-2-like module of PAXT. (7/x)
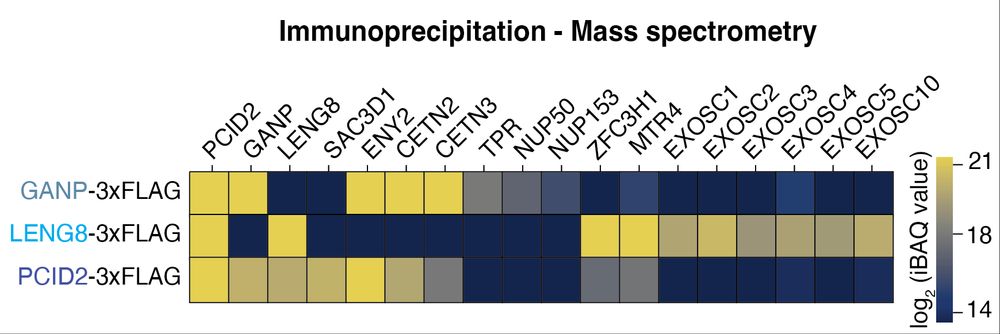
To our great surprise we found that LENG8-PS links to the ′Poly(A) tail exosome targeting (PAXT)’ connection, implicated in the degradation of polyadenylated nuclear RNA through the exosome. (6/x)
September 22, 2025 at 11:23 PM
To our great surprise we found that LENG8-PS links to the ′Poly(A) tail exosome targeting (PAXT)’ connection, implicated in the degradation of polyadenylated nuclear RNA through the exosome. (6/x)
Turns out they do! Both additional complexes bind UAP56 in vitro, and, like TREX-2, stimulate the release of RNA from UAP56. Seems counter-productive for mRNA export. So, what is their function? (5/x)

September 22, 2025 at 11:23 PM
Turns out they do! Both additional complexes bind UAP56 in vitro, and, like TREX-2, stimulate the release of RNA from UAP56. Seems counter-productive for mRNA export. So, what is their function? (5/x)
Most eukaryotes harbor two additional, less characterized TREX-2 like complexes: LENG8-PS and SAC3D1-PS, which interestingly differ in their subcellular localization. What is their function, and do they also act on UAP56? (4/x)

September 22, 2025 at 11:23 PM
Most eukaryotes harbor two additional, less characterized TREX-2 like complexes: LENG8-PS and SAC3D1-PS, which interestingly differ in their subcellular localization. What is their function, and do they also act on UAP56? (4/x)
In the final step of mRNA export UAP56 enables docking of the mRNP (mRNA+bound proteins) to TREX-2, which is anchored at the nuclear pore. TREX-2 in turn removes UAP56 from the mRNA, enabling the release of the mRNP from the nuclear pore and its shuttling to the cytoplasm. (3/x)

September 22, 2025 at 11:23 PM
In the final step of mRNA export UAP56 enables docking of the mRNP (mRNA+bound proteins) to TREX-2, which is anchored at the nuclear pore. TREX-2 in turn removes UAP56 from the mRNA, enabling the release of the mRNP from the nuclear pore and its shuttling to the cytoplasm. (3/x)
We previously suggested a model for a core mRNA nuclear export pathway, which centers on the action of the RNA clampase UAP56: www.biorxiv.org/content/10.1... (2/x)
September 22, 2025 at 11:23 PM
We previously suggested a model for a core mRNA nuclear export pathway, which centers on the action of the RNA clampase UAP56: www.biorxiv.org/content/10.1... (2/x)