




Juan Carlos De la Concepcion
@delaconcepcionjc.bsky.social
Plants, Pathogens and Procrastination
This "protein complex scape" represents an unexplored mechanism in which a subunit from a molecular complex can diversify and be co-opted into completely novel pathways, without interfering with the ancestral function of the complex.

October 31, 2025 at 1:57 PM
This "protein complex scape" represents an unexplored mechanism in which a subunit from a molecular complex can diversify and be co-opted into completely novel pathways, without interfering with the ancestral function of the complex.
Our data suggest that, during evolution, plant Exo70s accumulated negative charges at their N-termini, resulting on dissociation from the exocyst. This allowed Exo70s to break free from evolutionary constrains and interference, permitting their expansion and diversification

October 31, 2025 at 1:57 PM
Our data suggest that, during evolution, plant Exo70s accumulated negative charges at their N-termini, resulting on dissociation from the exocyst. This allowed Exo70s to break free from evolutionary constrains and interference, permitting their expansion and diversification
@josejulian.bsky.social validated the relation between net charge and Exo70 localization (a proxy for association to the exocyst) in Arabidopsis. AtExo70s with negative charge had a diffused location while those with neutral or positive localized at the cell plate!

October 31, 2025 at 1:57 PM
@josejulian.bsky.social validated the relation between net charge and Exo70 localization (a proxy for association to the exocyst) in Arabidopsis. AtExo70s with negative charge had a diffused location while those with neutral or positive localized at the cell plate!
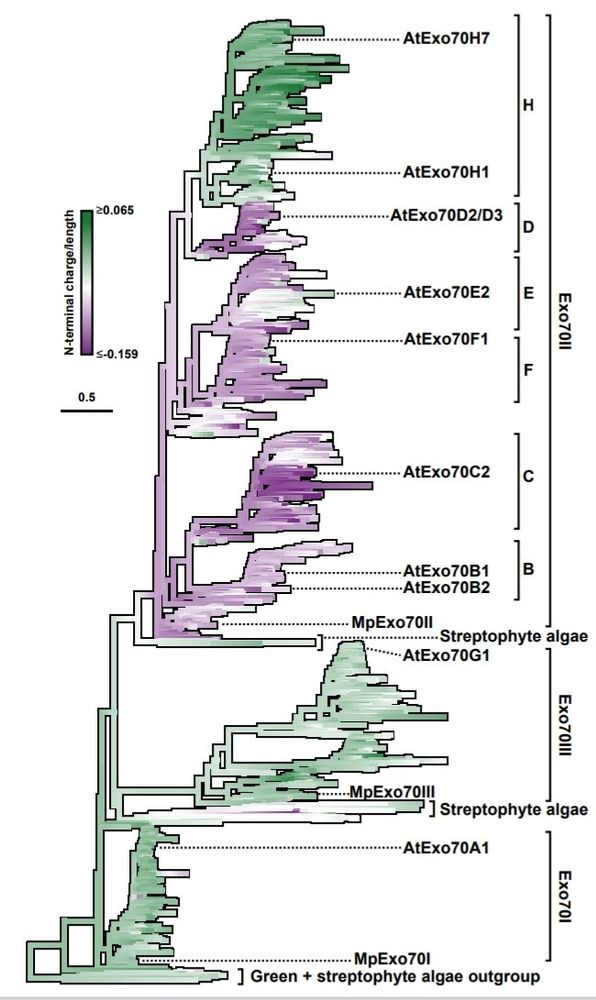
What were the implications of the N-terminal charge in Exo70 evolution? We did an ancestral gene reconstruction of the Exo70 N-termini and calculate the overall charge. This unvovered a switch towards negative charge that predated the emergence and expansion of Exo70 clade II 🤯
October 31, 2025 at 1:57 PM
What were the implications of the N-terminal charge in Exo70 evolution? We did an ancestral gene reconstruction of the Exo70 N-termini and calculate the overall charge. This unvovered a switch towards negative charge that predated the emergence and expansion of Exo70 clade II 🤯
And when we replaced MpExo70II Glu residues by Arg to neutralize the negative charge, this mutant now relocalized to the cell plate 😮. So the net charge at the N-terminus defines Exo70 association with the exocyst and the protein's localization ✅

October 31, 2025 at 1:57 PM
And when we replaced MpExo70II Glu residues by Arg to neutralize the negative charge, this mutant now relocalized to the cell plate 😮. So the net charge at the N-terminus defines Exo70 association with the exocyst and the protein's localization ✅
Is overall net charge underpinning the differences in Exo70 localization and exocyst association? We replaced MpExo70I residues at the N-terminus by Glu to simulate the charge of MpExo70II and this mutant now got a diffuse location...

October 31, 2025 at 1:57 PM
Is overall net charge underpinning the differences in Exo70 localization and exocyst association? We replaced MpExo70I residues at the N-terminus by Glu to simulate the charge of MpExo70II and this mutant now got a diffuse location...
This leads to an overall negative charge at the N-terminus of Exo70s from clade II compared to clade I and III

October 31, 2025 at 1:57 PM
This leads to an overall negative charge at the N-terminus of Exo70s from clade II compared to clade I and III
Linear discriminant analysis revealed the source of the differences at Exo70II N-termini is an accumulation of negatively charged aminoacid (Asp or Glu)

October 31, 2025 at 1:57 PM
Linear discriminant analysis revealed the source of the differences at Exo70II N-termini is an accumulation of negatively charged aminoacid (Asp or Glu)
So, what are the Exo70 differences underpinning these differential interaction? With Nick Irwin we looked at the diversity in the plant Exo70 family and found that the N-termini from clade II are the most variable

October 31, 2025 at 1:57 PM
So, what are the Exo70 differences underpinning these differential interaction? With Nick Irwin we looked at the diversity in the plant Exo70 family and found that the N-termini from clade II are the most variable
Overexpression of Exo70 N-terminal domain recapitulated the phenotypes we got when expressing Exo70 lipid binding mutants, confirming the importance of this domain in Exo70 localization, interactions and function

October 31, 2025 at 1:57 PM
Overexpression of Exo70 N-terminal domain recapitulated the phenotypes we got when expressing Exo70 lipid binding mutants, confirming the importance of this domain in Exo70 localization, interactions and function
Mainly because the Exo70 N-terminal domain underpins the interaction with the core component Exo84 and, therefore, association to the exocyst

October 31, 2025 at 1:57 PM
Mainly because the Exo70 N-terminal domain underpins the interaction with the core component Exo84 and, therefore, association to the exocyst
This is because the chimeras also swapped their ability to associate with the exocyst complex

October 31, 2025 at 1:57 PM
This is because the chimeras also swapped their ability to associate with the exocyst complex
Exchanging the N-terminal region of Exo70I and Exo70II swapped their cellular localization

October 31, 2025 at 1:57 PM
Exchanging the N-terminal region of Exo70I and Exo70II swapped their cellular localization
What are the molecular determinants of these differences? Taking advantage of the structural similarity between Exo70I and Exo70II, we designed chimeric protein swapping structurally equivalent regions

October 31, 2025 at 1:57 PM
What are the molecular determinants of these differences? Taking advantage of the structural similarity between Exo70I and Exo70II, we designed chimeric protein swapping structurally equivalent regions
We confirmed this using Arabidopsis, in which Marchantia Exo70I but not Exo70II complemented exocyst defects caused by the mutation of AtExo70A1. This all suggested that Exo70II has a different function from canonical exocytosis.

October 31, 2025 at 1:57 PM
We confirmed this using Arabidopsis, in which Marchantia Exo70I but not Exo70II complemented exocyst defects caused by the mutation of AtExo70A1. This all suggested that Exo70II has a different function from canonical exocytosis.
We could not obtain CRISPR mutants for any Marchantia Exo70 (likely essential genes). To circunvent this we designed lipid binding mutants that could act as dominant negative upon conditional expression. Interestingly, Exo70I and Exo70II mutants led to different phenotypes!

October 31, 2025 at 1:57 PM
We could not obtain CRISPR mutants for any Marchantia Exo70 (likely essential genes). To circunvent this we designed lipid binding mutants that could act as dominant negative upon conditional expression. Interestingly, Exo70I and Exo70II mutants led to different phenotypes!
We also find differences in ability to bind to phospholipids between Exo70I and Exo70II, further indicating a possible functional diversification

October 31, 2025 at 1:57 PM
We also find differences in ability to bind to phospholipids between Exo70I and Exo70II, further indicating a possible functional diversification
This reduced association is because, contrary to ExoI and ExoIII, ExoII does not interact with the core exocyst component Exo84

October 31, 2025 at 1:57 PM
This reduced association is because, contrary to ExoI and ExoIII, ExoII does not interact with the core exocyst component Exo84
... and even more interesting, it turned out that ExoII has a reduced association to the exocyst complex. That explain why this protein is diffused and not co-localize with the exocyst! 🧐

October 31, 2025 at 1:57 PM
... and even more interesting, it turned out that ExoII has a reduced association to the exocyst complex. That explain why this protein is diffused and not co-localize with the exocyst! 🧐
We set out to understand what are the differences between Exo70 paralogs using IP-MS and found that each Exo70 has its own interactome...

October 31, 2025 at 1:57 PM
We set out to understand what are the differences between Exo70 paralogs using IP-MS and found that each Exo70 has its own interactome...
Using confocal microscopy,
we found that, while Exo70I and Exo70III co-localize with the other exocyst components (SEC6 and Exo84) at the cell plate, ExoII has a diffused celullar pattern 😮
we found that, while Exo70I and Exo70III co-localize with the other exocyst components (SEC6 and Exo84) at the cell plate, ExoII has a diffused celullar pattern 😮

October 31, 2025 at 1:57 PM
Using confocal microscopy,
we found that, while Exo70I and Exo70III co-localize with the other exocyst components (SEC6 and Exo84) at the cell plate, ExoII has a diffused celullar pattern 😮
we found that, while Exo70I and Exo70III co-localize with the other exocyst components (SEC6 and Exo84) at the cell plate, ExoII has a diffused celullar pattern 😮
To investigate how Exo70s underwent extreme diversification without compromising the role of complex, we used the powerful 🌱 model #Marchantia, which has only 3 Exo70 paralogs (Exo70I, II and III).

October 31, 2025 at 1:57 PM
To investigate how Exo70s underwent extreme diversification without compromising the role of complex, we used the powerful 🌱 model #Marchantia, which has only 3 Exo70 paralogs (Exo70I, II and III).
In animals and fungi, you normally find a single copy of each subunit of the complex. In plants, the Exo70 subunit has dramatically expanded up to dozens of paralogs! 💥 And multiple paralogs have been linked to different functions! How is this possible? 🤔

October 31, 2025 at 1:57 PM
In animals and fungi, you normally find a single copy of each subunit of the complex. In plants, the Exo70 subunit has dramatically expanded up to dozens of paralogs! 💥 And multiple paralogs have been linked to different functions! How is this possible? 🤔
In summary, we show how alleles of the helper NLR Pik-2 have differentially co-evolved with their matching sensor NLR Pik-1. Pikm-2 acquired an Asp to Glu mutation that ensured an efficient response to a larger subset of AVR-Pik effectors but, as a consequence, became incompatible with Pikp-1.

April 2, 2025 at 10:20 AM
In summary, we show how alleles of the helper NLR Pik-2 have differentially co-evolved with their matching sensor NLR Pik-1. Pikm-2 acquired an Asp to Glu mutation that ensured an efficient response to a larger subset of AVR-Pik effectors but, as a consequence, became incompatible with Pikp-1.
Replacing the Glu polymorphism by the ancestral Gly residue abrogated the constitutive NLR activation, confirming this phenotype is a byproduct of Pikm-2 evolution towards a fine-tuned response to effectors.

April 2, 2025 at 10:20 AM
Replacing the Glu polymorphism by the ancestral Gly residue abrogated the constitutive NLR activation, confirming this phenotype is a byproduct of Pikm-2 evolution towards a fine-tuned response to effectors.