



Madison Chapel
@chapelmadison.bsky.social
MSc, bioinformatics (UBC)
Head full of yarn scraps, bloodstream full of bubble tea
Head full of yarn scraps, bloodstream full of bubble tea
11/ Our simulations suggest that, rather than being a direct target of selection, GRN complexity may arise as a byproduct of other evolutionary processes. And to close things out, here’s an animation of a GRN evolving. Watch how fitness and complexity change as the population evolves ! 🎥
September 3, 2025 at 7:25 PM
11/ Our simulations suggest that, rather than being a direct target of selection, GRN complexity may arise as a byproduct of other evolutionary processes. And to close things out, here’s an animation of a GRN evolving. Watch how fitness and complexity change as the population evolves ! 🎥
10/ We saw that complexity emerged more rapidly under changing environmental conditions. For recombining populations, this effect was particularly pronounced.

September 3, 2025 at 7:25 PM
10/ We saw that complexity emerged more rapidly under changing environmental conditions. For recombining populations, this effect was particularly pronounced.
9/ We simulated changing environments by shifting the expression goal for a subset of genes. You can see fitness decrease sharply as the environment changes, followed by recovery as populations adapt.

September 3, 2025 at 7:25 PM
9/ We simulated changing environments by shifting the expression goal for a subset of genes. You can see fitness decrease sharply as the environment changes, followed by recovery as populations adapt.
7/ So what did recombination do? First off, recombining populations were consistently more 𝗿𝗼𝗯𝘂𝘀𝘁 than non-recombining ones – they were better at maintaining an expression profile after mutational perturbation. This matches observations from many previous studies!

September 3, 2025 at 7:25 PM
7/ So what did recombination do? First off, recombining populations were consistently more 𝗿𝗼𝗯𝘂𝘀𝘁 than non-recombining ones – they were better at maintaining an expression profile after mutational perturbation. This matches observations from many previous studies!
5/ Surprisingly, recombination didn’t change the final complexity. Neither mutation rate nor initial binding affinity mattered, either. Given enough time, everything converged to ‘random’ GRN complexity; the same level seen in populations evolved 𝘄𝗶𝘁𝗵𝗼𝘂𝘁 𝗮𝗻𝘆 𝘀𝗲𝗹𝗲𝗰𝘁𝗶𝗼𝗻.

September 3, 2025 at 7:25 PM
5/ Surprisingly, recombination didn’t change the final complexity. Neither mutation rate nor initial binding affinity mattered, either. Given enough time, everything converged to ‘random’ GRN complexity; the same level seen in populations evolved 𝘄𝗶𝘁𝗵𝗼𝘂𝘁 𝗮𝗻𝘆 𝘀𝗲𝗹𝗲𝗰𝘁𝗶𝗼𝗻.
4/ To investigate this, we built a biochemically-inspired GRN model. TF affinities and concentrations determine gene expression. The closer expression levels are to a specific goal, the more fit a GRN is. Using this model, we simulated 1 million generations of evolution.

September 3, 2025 at 7:25 PM
4/ To investigate this, we built a biochemically-inspired GRN model. TF affinities and concentrations determine gene expression. The closer expression levels are to a specific goal, the more fit a GRN is. Using this model, we simulated 1 million generations of evolution.
3/ Here’s our idea: complexity in eukaryotic GRNs may have evolved in response to recombination. In simple GRNs, recombination could introduce differently acting transcription factors (TFs) that alter gene expression. In complex GRNs, expression would be buffered by additional TFs

September 3, 2025 at 7:25 PM
3/ Here’s our idea: complexity in eukaryotic GRNs may have evolved in response to recombination. In simple GRNs, recombination could introduce differently acting transcription factors (TFs) that alter gene expression. In complex GRNs, expression would be buffered by additional TFs
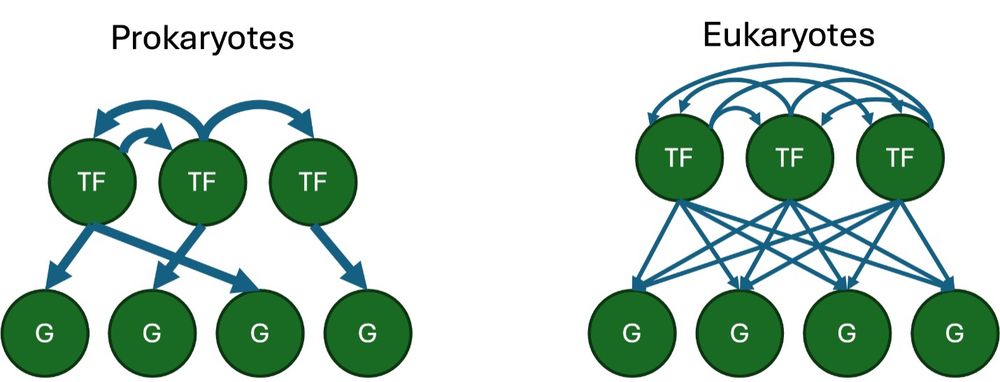
2/ Prokaryotic GRNs are relatively simple – there’s specific binding between TFs and their target sites. But in complex eukaryotic GRNs, TFs recognize identical sites throughout the genome, and expression of any one gene depends on multiple TFs binding in combination.
September 3, 2025 at 7:25 PM
2/ Prokaryotic GRNs are relatively simple – there’s specific binding between TFs and their target sites. But in complex eukaryotic GRNs, TFs recognize identical sites throughout the genome, and expression of any one gene depends on multiple TFs binding in combination.
10/ We saw that complexity emerged more rapidly under changing environmental conditions. For recombining populations, this effect was particularly pronounced.
September 3, 2025 at 7:15 PM
10/ We saw that complexity emerged more rapidly under changing environmental conditions. For recombining populations, this effect was particularly pronounced.
9/ We simulated changing environments by shifting the expression goal for a subset of genes. You can see fitness decrease sharply as the environment changes, followed by recovery as populations adapt.
September 3, 2025 at 7:15 PM
9/ We simulated changing environments by shifting the expression goal for a subset of genes. You can see fitness decrease sharply as the environment changes, followed by recovery as populations adapt.
7/ Polygenic background also changes how selection acts! More alleles contributing to a trait widens the genetic risk distribution, increasing selective pressure as individuals are pushed to extremes. Highly polygenic traits lead to smaller, more uniform effect sizes.

January 8, 2025 at 8:24 PM
7/ Polygenic background also changes how selection acts! More alleles contributing to a trait widens the genetic risk distribution, increasing selective pressure as individuals are pushed to extremes. Highly polygenic traits lead to smaller, more uniform effect sizes.
4/ Disease prevalence scales non-linearly with disease liability. A 1-unit increase in PGS leads to very different changes in disease prevalence depending on whether it occurs in a low-risk (green) or high-risk (red) genetic background. Same variant, totally different outcomes!

January 8, 2025 at 8:24 PM
4/ Disease prevalence scales non-linearly with disease liability. A 1-unit increase in PGS leads to very different changes in disease prevalence depending on whether it occurs in a low-risk (green) or high-risk (red) genetic background. Same variant, totally different outcomes!